
En un artículo anterior analizamos las teorías de Edelman y Tononi sobre la base neural de la consciencia primaria, que es una capacidad para representar perceptualmente el mundo exterior, reconocer sus cualidades y reaccionar ante ellas. Un paso más allá de la consciencia primaria, que sólo los humanos y los mamíferos superiores han dado, es la capacidad llamada consciencia autorreflexiva, o consciencia de uno mismo como testigo de la consciencia primaria. Un paso necesario para alcanzar una consciencia autorreflexiva, y todo lo que ella permite (la evolución cultural, por ejemplo) es la capacidad de reconocer el propio cuerpo y el cuerpo de otros como similares, e imitar lo que los cuerpos de otros saben hacer.
Estudios de Meltzoff y Moore (1997), a los que pertenece la imagen que encabeza este artículo), y otros estudios, defienden que los bebés de sólo tres semanas pueden imitar la protrusión de la lengua de un adulto; y que con dos meses imitan gestos de un adulto como los siguientes: protrusión de la lengua, protrusión de los labios, apertura de la boca, gestos con las manos, movimientos de la cabeza, parpadeo de los ojos, movimientos de las mejillas y las cejas, y componentes de las expresiones emocionales.
En un bebé que imita la expresión facial de un adulto, ¿Cómo hace el cerebro del niño para hacer coincidir un gesto propio que siente, pero no puede ver, con el gesto del adulto, que ve pero no puede sentir? Como explican Meltzoff y Moore (1997): “Teorías clásicas como la de Piaget (1962) respondieron esta pregunta a través de experiencias de aprendizaje con espejos y exploración táctil por el bebé de las caras propias y ajenas. Los espejos hicieron visible lo invisible, representando el propio cuerpo y el del otro en términos visuales. La exploración táctil de rostros se representa a sí mismo y al otro en términos tangibles. En los últimos 20 años, el trabajo empírico de muchos laboratorios ha revelado que bebés demasiado pequeños para haber aprendido de esa manera imitan sin embargo los gestos faciales.”
La respuesta alternativa de Meltzoff y Moore (1997) es que los bebés de esa edad han construido ya en su cerebro un mapa conceptual intermodal (que agrupa información de distintos sentidos) del propio cuerpo y a partir de él deducen los actos motores posibles. Según estos autores, el bebé nace con un mapa propioceptivo-motor de su cuerpo ya desarrollado, lo cual le permite identificar rápidamente las imágenes visuales de las partes corporales de otros y crear un mapa corporal visual isomorfo con el anterior, y finalmente, traducir imágenes del movimiento de miembros corporales a actos motores.
En un curioso experimento, cuando el adulto sacaba la lengua diagonalmente de la boca, el 70% de los bebés de un mes y medio de edad sacaron la lengua y giraron la cabeza 30 grados, alcanzando una orientación de lengua similar, aunque no exactamente el mismo gesto. Es como si su comportamiento estuviera dirigido a un objetivo.
Cuando un recién nacido ve una protrusión de lengua, su primera reacción es aquietar todas las partes del cuerpo menos la lengua, que se activa, sin hacer necesariamente una protrusión de inmediato, sino que la agita, la mueve o la levanta dentro de la cavidad bucal. Del mismo modo, cuando se les muestra una protrusión de los labios, lo primero que hacen es presionar fuertemente un labio contra el otro, antes de intentar la imitación. Es como si aislaran primero qué parte del cuerpo van a mover, antes de decidir cómo hacerlo. La identificación la realiza aparentemente el bebé sobre el conjunto de los órganos, independientemente de que pertenezcan a él o a otro.
Hay evidencias de que el bebé recién nacido reconoce un conjunto delimitado de órganos en función de su forma (`órganos como formas ‘). Varios autores han observado que los recién nacidos prefieren las caras a otros patrones sensoriales. A través de la evolución, la estructura de una cara podría haberse convertido en una unidad perceptiva distintiva a la que se responde ya de forma innata. Puede ser que el contexto perceptual de una cara ayude al bebé a identificar el conjunto, cuyas partes componentes pueden luego analizar. Algunas investigaciones sugieren que los órganos faciales pueden ser perceptualmente identificables por los recién nacidos humanos, aunque no hay acuerdo completo sobre el tema. Además, los datos neurofisiológicos muestran que las imágenes visuales de órganos particulares, especialmente partes de la cara y manos en los monos, activan sitios específicos del cerebro. De modo que hay órganos específicos que podrían estar representados ya al nacer en el cerebro del bebé.
La psicología cognitiva ha desarrollado modelos sobre cómo podría conceptualizarse la percepción visual de un cuerpo animal. Los grupos neuronales especialistas detectarían pautas que, conjuntamente, serían etiquetadas en forma de concepto no-verbal por algún área semántica del córtex, especialista en integrar y etiquetar (no verbalmente) pautas sensoriales de todos los especialistas de la percepción visual. Una etiqueta no verbal equivalente a un concepto abstracto del tipo, por ejemplo, de “pierna delgada”.
La construcción de objetos por la percepción visual
Desde el momento en que la retina convierte fotones luminosos en impulsos neuronales deja de tener sentido hablar de la información visual como si fuera una imagen. Se trata más bien de descripciones simbólicas que tratan de dar forma (in-formar) la percepción visual de forma que sea manipulable o significativa para el cuerpo. Esto lo realizan unas 30 áreas neuronales especialistas del sistema visual de los humanos (los herbívoros y otros mamíferos superiores tienen unas 12) que extraen rasgos significativos de la percepción. Áreas especializadas en reconocimiento del color, de diferentes formas, de los bordes y contrastes, del movimiento, de si se trata de una cara, etc. Las sinapsis neuronales que dejan la retina son unas 100 veces menos numerosas que las que tienen lugar en la propia retina, por lo que podríamos decir que ya desde la retina se empieza a producir una condensación de la imagen original en una serie de rasgos significativos que son enviados hacia las áreas visuales del cerebro para ser elaborados y depurados más aún.
Este proceso cerebral es activo e incluye prejuicios sobre lo que puede ser observado. Por ejemplo, en la figura siguiente parece claro cuáles son los círculos convexos y cuáles los cóncavos.

Sin embargo, si ponemos esa figura horizontalmente, nuestra percepción visual oscila entre verlos de esa manera o de la manera complementaria.

Lo más verosímil es imaginar que algún área especializada del cerebro está pre-suponiendo que, en la primera imagen, la luz ilumina a los objetos desde arriba (la iluminación desde abajo no ocurre en la naturaleza); en la segunda imagen, en cambio, no tiene ninguna predilección por suponer que la luz venga de la derecha o de la izquierda, y alterna entre ambos presupuestos cada cierto tiempo. Para ser más exactos, la suposición de nuestro cerebro es que la luz viene desde encima de la propia cabeza, y eso se ve claramente si contemplamos la última de las figuras poniéndonos nosotros horizontales, no la figura. Observaremos entonces que la ambigüedad desaparece, pues vemos irremisiblemente todas las figuras como si estuvieran iluminadas desde encima de nuestras cabezas (Ramachandran 2011, Cap. 2).
Otra característica notable del sistema visual es que a medida que subimos hacia las áreas especializadas del córtex cerebral desde la retina, pasando por el núcleo geniculado lateral del tálamo, las conexiones que van desde etapas superiores hacia etapas anteriores son mucho más numerosas que las que ascienden. Ello sugiere una dinámica bayesiana del procesamiento de la información (véase post 1 **), en la que el córtex superior está enviando alucinaciones todo el tiempo hacia abajo, hasta que la información que asciende desde la retina deja de ser incoherente con la última alucinación propuesta, y a ésta la seleccionamos y etiquetamos finalmente como el concepto percibido (Ramachandran 2011, Cap. 2; Dennett 2017).
Desde el punto de vista neurológico, hay dos caminos desde la retina hasta el área visual V1 del lóbulo parietal (área 17 de Brodmann). Un camino (que Ramachandran llama el “camino antiguo”) pasa por el colículo superior y acaba en V1.

El otro camino (el “camino nuevo”) pasa por el núcleo geniculado lateral del tálamo, desde allí se dirige hasta V1, en el lóbulo occipital, y allí se bifurca en dos ramas: la rama del “donde” (también llamada del “como”) se dirige hacia el lóbulo parietal, y la rama del “qué” se dirige hacia el área 21 de Broadman, en el lóbulo temporal.

La primera rama está especializada en las relaciones espaciales entre los objetos identificados; la segunda, en las relaciones entre las propiedades encontradas dentro de cada objeto y lo que significan para uno, lo cual permitirá etiquetar posteriormente al objeto dentro de una categoría lingüística.

El camino “antiguo” puede dirigir la mano de una persona hacia un objeto en el campo visual de forma automática sin que la persona sea consciente de que está viendo el objeto. El camino “nuevo” permite, sin embargo, la experiencia consciente de estar viendo un objeto (Ramachandran 2011, Cap. 2).
La primera estructura de la ruta del “qué”, el giro fusiforme, parece clasificar el objeto visual en distintas categorías conceptuales (diferencia un pájaro de un humano, o un cuchillo de un piano) pero con poca elaboración sobre el significado del concepto para el observador. Pero cuando esta ruta se adentra más profundamente en los lóbulos temporales, evoca no sólo la categoría cruda sino un conjunto difuso de recuerdos asociados y hechos y usos del objeto, es decir, su significado para el observador. Esta recuperación semántica implica la activación de conexiones con otras áreas del lóbulo temporal, con el área lingüística de Wernicke y con el lóbulo parietal inferior, que está especializado en poner etiquetas lingüísticas (nombrar), escribir, y en la aritmética. Tras el apoyo de estas regiones, los mensajes se transmiten a la amígdala, que se encuentra incrustada en la punta frontal de los lóbulos temporales, donde evocan sentimientos relacionados con lo que (o quien) estás viendo (Ramachandran, 2011, Cap. 2).
Sin embargo, si el objeto visual tiene una alta saliencia (es inhabitual) el camino 2 no sigue la ruta habitual, sino que abrevia desde el giro fusiforme, pasando por surco temporal superior, hasta entrar directamente en la amígdala. De esta manera, evita pasar por las áreas que enriquecen el objeto visual con significaciones, y entra rápidamente en la amígdala, en el corazón emocional del cerebro, el sistema límbico. Este camino rápido evolucionó probablemente para promover reacciones rápidas ante escenas inhabituales, ya sean innatas o aprendidas. La amígdala trabaja junto con recuerdos que ayudan a calibrar la importancia emocional de la escena visual. Si la respuesta emocional de la amígdala es intensa (con emociones de hambre, miedo, odio o lujuria, por ejemplo), influye en el hipotálamo, que induce una liberación de las hormonas necesarias para preparar el sistema nervioso autónomo para una próxima acción (alimentarse, luchar, huir, cortejar, etc.). Esto incluye aumento del ritmo cardíaco, respiración rápida y superficial, y aumento de la transpiración (que aumenta la refrigeración del sistema motor). Estas reacciones fisiológicas que van asociadas a los sentimientos son las que aprovechan los detectores de mentiras para saber si la afirmación de alguien es congruente o no con su reacción emocional. La amígdala está conectada también con los lóbulos frontales, que añaden matices a estas emociones primarias (v.g. arrogancia, prudencia, orgullo, admiración, magnanimidad, etc.).

Cote transversal y vista lateral del cerebro, para ilustrar la posición del sistema límbico. De: http://www.facmed.unam.mx/Libro-NeuroFisio/06-SistemaNervioso/CNS-Overview/SistNervioso.html
El trabajo https://webs.ucm.es/info/pslogica/mente/cap3.htm resume muy bien las tres etapas principales del proceso de construcción conceptual de un objeto perceptual. (1) En un primer nivel de procesamiento neuronal, se hace un análisis del objeto visual que pretende detectar agrupamientos importantes de la imagen, los bordes o perfil de la misma, y los elementos constitutivos de la imagen desde el punto de vista del plano. Produce una representación primaria, o Bosquejo Primario. Sería algo similar a la detección de bordes que realizan algunos programas gráficos de ordenador.
(2) El segundo nivel de procesamiento, que toma como entrada el bosquejo primario, producirá un bosquejo en 2 dimensiones al que se ha añadido algo de información cruzada procedente de los dos ojos, que permite etiquetar qué objetos están delante de cuáles, pero aún centrado en el observador. Este proceso tiende a construir la superficie de la imagen, añadiéndola textura, profundidad y orientación, captando movimiento y color. Podría ser algo similar a la información que proporciona el cuadro siguiente.

(3) El tercer nivel de procesamiento tiene como propósito lograr una representación en tres dimensiones que sea útil para el reconocimiento. Exige pasar a un sistema de coordenadas centrado en el objeto y no en el observador. También, incluir formas primitivas de diferentes tamaños en una organización modular que distinga la imagen como un todo con partes y finalmente, incluir esquemas de volúmenes que expliciten el espacio ocupado por los objetos que se supone componen la escena y no sólo sus superficies directamente visibles.
Si, por ejemplo, el nivel (2) ha obtenido la imagen de un burrito (a en la figura siguiente), la imagen es suavizada y dividida en secciones cóncavas y convexas (b). Después se identifican puntos de gran segmentación (c) que son conectados entre sí (d). A partir de ello, se definen los ejes componentes que aparecen en (e); y estos grupos se conectan como partes a un todo en (f) (figura siguiente). Este proceso de segmentación e identificación de las partes del objeto es clave para la capacidad de distinguir un objeto, incluso cuando una parte del mismo aparece ocultado por otro objeto que parece estar delante.
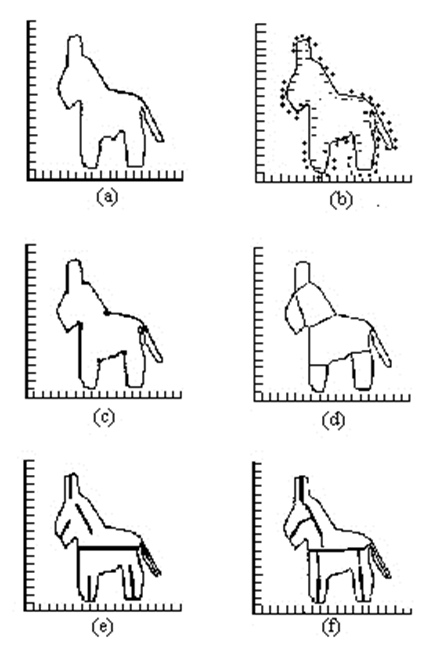
Proceso de identificación de las partes de un objeto. De: https://webs.ucm.es/info/pslogica/mente/cap3.htm
Los sistemas neuronales especializados en asignar a las imágenes del nivel 2 un fondo, y por lo tanto un volumen espacial, están categorizando las impresiones visuales básicas organizándolas en el espacio, algo que también hace el bebé dentro del útero cuando va formando el mapa sensorio-motor de su cuerpo, también organizado espacialmente. Esta pulsión, programada prácticamente en nuestra fisiología de inicio, a conceptualizar las sensaciones (tanto táctiles como visuales) en forma espacial, era para Kant (y para su discípulo Schopenhauer) una de los más importantes aprioris de la apercepción trascendental, aquellos que nos permiten percibir un mundo, entenderlo y actuar en él. Los grupos neuronales que realizan esta importante transformación podrían ser también la clave de que un niño de mes y medio de edad haya aprendido en ese breve tiempo a asociar a las imágenes que percibe de cuerpos humanos, animales y objetos, conceptos espaciales análogos a los que inconscientemente está usando para concebir su propio cuerpo. Por ejemplo, diferencia la parte frontal de un cuerpo u objeto externo de su parte trasera; y va creando rápidamente conceptos espaciales para ellos, su entorno espacial y la relación entre ellos y nuestro cuerpo, conceptos que más tarde serán etiquetados lingüísticamente como cercano-lejano, delante-detrás, cabeza-piés, interior-exterior, sobre-debajo… Es posible que esa capacidad de identificar volúmenes y relaciones topológicas permita al bebé de mes y medio identificar conceptualmente los principales órganos del cuerpo de los adultos que lo rodean como los órganos análogos al que su mapa sensorio-motor le hace sentir en su propio cuerpo. La propia observación visual de sus miembros probablemente refuerza esa asociación, a lo largo de los primeros meses.
El blog citado sigue la explicación de D. Marr (1985) sobre cómo puede estar trabajando a nivel neurológico el sistema visual. El paso siguiente sería la organización de la información sobre la forma en un modelo 3-D, y con un sistema de coordenadas centrado en el objeto. Un esquema del proceso sería el siguiente.

Para cada región significativa se construye un modelo tridimensional que se agrupan modularmente formando un todo. A cada módulo se la asigna también un sistema de coordenadas propio, pero bajo el entendido de que se trata de partes de una misma escena visual. Esto facilitará las tareas de reconocimiento, al permitir comparar la representación construida con el conocimiento conceptual previo que tenemos de los elementos de mundo. Por ejemplo, se identifica mejor un dedo si se relaciona con la mano de la que es parte, y así sucesivamente.
Hasta aquí https://webs.ucm.es/info/pslogica/mente/cap3.htm
La distinción entre percepción y acción no es absoluta para el cerebro, como lo demuestran las neuronas canónicas de los lóbulos frontales, un tipo de neuronas espejo que según Ramachandran (2011, Cap. 2) se disparan en acciones específicas como alcanzar una manzana o una ramita, pero también lo hacen cuando el sistema perceptivo identifica una manzana o una ramita. Es como si el concepto de agarrabilidad fuera añadido a los demás atributos del percepto.
Por último, el proceso de percepción termina colocando un nombre a la representación construida. Este proceso, bastante desconocido aún, sería semejante a una búsqueda en un catálogo de esquemas de imágenes, junto con conexiones al sistema conceptual y lingüístico. Las tres áreas del lenguaje están especializadas en estas funciones: el área de Broca (en lóbulo frontal), el área de Wernicke (en lóbulo temporal) y el giro angular (cerca del fondo del lóbulo parietal). Lakoff y Johnson (1999) (véase Las metáforas y la construcción imaginaria de la realidad) han analizado, de forma muy convincente, los procesos metonímicos y metafóricos que permiten construir conceptos complejos a partir de los primeros conceptos perceptivos, del mapa multi-modal del propio cuerpo, y de las primeras interacciones motoras entre el cuerpo del niño y el entorno habitual.
Volvamos ahora a la asociación que hace el niño entre el mapa sensorio-motor de su cuerpo y el mapa conceptual, que construye durante sus primeros meses, de los cuerpos familiares que observa.
Imaginemos, por ejemplo, que el bebé ha convivido con perros desde el nacimiento, algo que ha ocurrido realmente en algunos casos entre los llamados niños salvajes. Si el cerebro del bebé ha construido un mapa conceptual del cuerpo visual del perro que tiene delante, ese mapa conceptual es probablemente asociado, mediante la observación del cuerpo propio y mediante mecanismos de contigüidad topológica, al mapa sensorio-motor que el sujeto humano se ha construido de su propio cuerpo.
Una vez establecida la asociación conceptual entre las partes visuales de un cuerpo de la familia del mío y las impresiones sensorio-motoras que tengo en las partes correspondientes de mi propio cuerpo, esa asociación podría utilizarse para proporcionar información en cualquiera de los dos sentidos: el inicio del movimiento (o una sensación táctil o propioceptiva) en una de mis extremidades activaría la imagen de esa clase de extremidad; y viceversa, la imagen visual de una extremidad (mía o de un cuerpo de la familia del mío) moviéndose (o tocando algo) activaría el concepto propioceptivo-motor correspondiente a esa extremidad. También facilita el aprendizaje de lo que se puede hacer con el propio cuerpo, a partir de la imitación de los otros familiares, de ahí que los niños salvajes que convivieron desde casi el nacimiento sólo con perros, acaben caminando a cuatro patas, rascándose como los perros, etc.
Las acciones motoras
Según Meltzoff y Moore (1997), este aprendizaje propioceptivo-motor se realizaría mediante lo que llaman balbuceo corporal: ya en el útero, el bebé prueba al azar distintas acciones motoras y asocia los estados finales (propioceptivos) de las posiciones relativas de los distintos órganos con esos actos motores (y sus movimientos asociados). Por ejemplo, tres estados finales de la lengua tras movimientos de menor a mayor longitud podrían conceptualizarse de un modo no verbal pero señalando situaciones como las que designamos lingüísticamente con expresiones del tipo: “lengua a los labios”, “lengua entre los labios” y “lengua más allá de los labios”. Los recién nacidos podrían haber desarrollado en el útero un mapeo grosero entre estados finales de los órganos (sentidos propioceptivamente) y los movimientos necesarios para conseguirlos.
Experimentos de otros autores no han podido confirmar, sin embargo, que las observaciones de Meltzoff y Moore (1997) en niños de sólo mes y medio correspondan exactamente a imitación. Concluyen más bien que el bebé aumenta su excitación al reconocer la cara de su madre o de su padre, y saca continuamente la lengua, independientemente de lo que haga el adulto (Sticking the tongue out: Early imitation in infants) . Algunos investigadores sugieren que los bebés no tienen una predisposición innata a imitar, sino que aprenden a imitar siendo imitados. Los padres imitan continuamente al bebé cuando observan un gesto en él. Por ejemplo, cuando el bebé saca la lengua o produce un sonido bucal («ah!»), es muy probable que el adulto que está en frente sacque su lengua o imite el sonido («ah!»). Este comportamiento imitativo de los padres permitiría al bebé hacer las primeras asociaciones entre actos motores propios y el comportamientos visual de los demás.
Por tanto, el aprendizaje de los mapas corporales y la relación entre ellos podría no ser tan rápido como lo que sugieren Meltzoff y Moore (1997). Estos autores están de acuerdo en que los bebés no saben al nacer qué actos musculares consiguen reproducir un gesto particular de un órgano, y que esto se conseguiría por aprendizaje de prueba y error. Por tanto, la polémica está en (i) confirmar si el bebé nace ya con un mapa sensorio-motor de su cuerpo lo suficientemente detallado, y (ii) desarrolla en sólo mes y medio un mapa visual conceptual de los cuerpos familiares, de modo que a esa edad puede imitar gestos; o bien alguno (o ambos) de estos dos procesos es más prolongado en el tiempo de lo que sugieren Meltzoff y Moore.
Sí parece razonable suponer que los bebés interpretan el acto adulto que ven analizando las relaciones entre órganos que exhibe ese acto. Las relaciones entre los órganos proporcionan el marco común en el que se registran los actos propios y ajenos. ‘Lengua a los labios’, que es una relación entre órganos, sería una descripción conceptual (no verbal) que es isomorfa tanto con una percepción de un cuerpo observado como con unas sensaciones propioceptivas y táctiles en el cuerpo propio. Por lo tanto, las relaciones entre órganos harían que el acto facial observado pero no sentido del adulto sea comparable con el acto facial sentido pero invisible del niño.
Los experimentos muestran además que los bebés no se limitan a imitar actos bien practicados que pueden ser imitados en el primer intento. Lo que hacen aparentemente es intentar imitar el gesto, no quedan satisfechos muchas veces con el resultado motor inicial, y corrigen sus movimientos a lo largo del tiempo. Parecen intentar un buen ajuste de las relaciones entre órganos más que reproducir bien un acto motor ya ensayado. Por ejemplo, al ver al adulto sacar su lengua diagonalmente hacia su derecha, y recogerla también diagonalmente, muchos bebés de seis semanas mueven su lengua directamente a su derecha y la recogen; luego corrigen y la sacan en línea recta y luego la mueven a la derecha, recogiéndola diagonalmente; luego la sacan en línea recta y la recogen en un semicírculo; luego la sacan y la recogen en línea recta; finalmente, algunos bebés consiguen sacarla diagonalmente y recogerla diagonalmente; otros, se conforman con sacarla en línea recta, luego la mueven perpendicularmente a la derecha, y la recogen siguiendo los mismos pasos pero a la inversa.
Al ver el gesto de un adulto, un sistema de interpretación visual en el cerebro del bebé interpretaría la relación entre los órganos del adulto antes y después del movimiento gestual, y compararía el estado final con la relación entre los órganos de su propio cuerpo. Al no encontrar coincidencia, intentaría un acto motor que aproximadamente lleve a una relación mejor entre sus órganos. Encontrar una coincidencia indicaría que el acto motor visto y el acto motor realizado por uno mismo son equivalentes. Según estos dos autores, este reconocimiento de la equivalencia de los actos es motivo de la aprensión de los infantes de que el otro es, en cierto sentido primitivo y no consciente, ‘como yo’.
Neuronas espejo, aprendizaje y teoría de la mente
Leslie (1997) piensa que el ser humano nace con unas predisposiciones para procesar la información que se han desarrollado por selección natural. Son estructuras incluídas en la fisiología del sistema nervioso, y las califica de «teorías» por cuanto son como formas a priori, empleando terminología kantiana, para representar y categorizar la realidad.
Se darían dos tipos de teorías: Una teoría de objetos (TOB, abreviatura de Theory of body) y una teoría de la mente (TOM, Theory of mind). La teoría de los objetos proporciona los esquemas básicos para conocer el mundo de objetos físicos, sus propiedades y relaciones. La existencia de regiones especializadas del córtex que construyen propiedades básicas de objetos visuales sugieren que sí que existen estos esquemas a priori. Lakoff opina que son estos a-priori los que proporcionan esa fuerte convicción que tenemos en el realismo ingenuo cuando nos relacionamos con objetos de tamaño parecido a nuestro cuerpo, que pueden ser abarcados visualmente de una sola vez y que pueden ser manipulados (Las metáforas y la construcción imaginaria de la realidad).
La teoría de la mente posibilitaría la comprensión del otro y las relaciones interpersonales. La estructura neural que le corresponde debió de surgir de un afortunado ensamblaje entre la capacidad cerebral de crar mapas sensorio-motores del propio cuerpo, la capacidad para asociar unívocamente ese mapa con los mapas conceptuales de la forma corporal que surge de la observación visual, y la curiosa capacidad de las neuronas espejo para excitarse cuando usamos un miembro y cuando lo vemos usar. Para funcionar, necesitaría activarse mediante el aprendizaje de que hay formas visuales externas que, efectivamente son topológicamente similares a los mapas sensorio-motores que el bebé desarrolla (ya sea dentro del útero o en los primeros meses de vida).
Catmur et al. (2008) entrenaron a adultos participantes para que imitaran los gestos del bebé de forma congruente en un grupo de ellos, y de forma no congruente en otro grupo distinto, y encontraron diferencias posteriores en las activaciones de los sistemas de neuronas espejo de los dos grupos de bebés. Ello sugiere que los sistemas de neuronas espejo son bastante flexibles para aprender nuevas conexiones, y no vendrían congénitamente pre-programados. O bien vienen pre-programados de nacimiento de una forma muy genérica, pero están abiertos al aprendizaje posterior. Pero Meltzoff y Marshall (2018) encuentran respuestas mayores cuando la imitación es congruente con el órgano propio movido en niños de pocos meses. Encontraron también que la excitación de las neuronas espejo del córtex somatotópico de los bebés cuando ven que alguien es tocado en la mano o en el pie, se percibe ya a los 7 meses de edad.
Para poder activarse como lo hacen, las neuronas (grupos neuronales) espejo deberían estar conectadas con un mapa corporal visual (intermediario entre la percepción visual y ellas mismas) que les informe de que la imagen observada corresponde al cuerpo de otro estirando su brazo derecho hacia una pelota; y el mapa conceptual del aspecto visual del cuerpo del bebé debería estar conectado con el mapa senso-motor del propio cuerpo del bebé que observa y reacciona.
Lo que harían las neuronas espejos pertenecientes al conjunto de neuronas involucradas en el movimiento de esa extremidad es activarse cuando se produce tal asociación entre visión de una acción por parte de una parte de un cuerpo externo y la parte correspondiente de mi propio cuerpo. Para evitar que esta activación lleve a un movimiento real de la propia extremidad, el cortex prefrontal envía señales inhibidoras a la zona motora cuando nota la activación de las neuronas espejo, aunque normalmente no es suficientemente rápida como para evitar una ligera iniciación del propio movimiento, casi imperceptible.
La activación de los grupos de neuronas espejo podría estar genéticamente programada en cuanto el bebé logra identificar fuera de él movimientos que están en el repertorio de los movimientos que él ha identificado como posibles para su cuerpo. Posteriormente, cuanto el bebé logre identificar contenidos mentales dentro de él (entre los 7 y 9 meses), como por ejemplo intenciones que anteceden a sus movimientos, estas mismas intenciones serán otorgadas automáticamente al otro, si le ve realizar el mismo movimiento. Todo ello, antes de que el niño aprenda a hablar.
Al ver el movimiento exterior de miembros corporales reconocibles, el córtex superior del bebé detectaría el output de las neuronas espejo y conceptualizaría la situación de un modo similar a esto: ”el mismo grupo neuronal se está activando en mi cerebro como lo haría si estuviera alcanzando una pelota; así que el otro debe estar intentando alcanzar esa pelota ahora”.
En un próximo post veremos que, entre los dos y seis meses, el bebé parece adquirir una consciencia creciente de que está físicamente separado de su madre y de otros cuerpos. Se forma un “sí mismo nuclear” en contraste con el otro nuclear. Veremos también cómo a partir de ese núcleo corporal logra generar una consciencia de sí mismo. El descubrimiento en el niño de que los demás cuerpos humanos son intencionales y mentales como él mismo, y que los contenidos emocionales y conscientes propios pueden ser compartidos con otros, se produce entre los 7 y los 9 meses. Al año el niño suele ser capaz de indicar a otro que le acerque un juguete. Pero esa “teoría de la mente” se va volviendo cada vez más compleja y detallada entre el año y los 5 años.
Se han encontrado neuronas espejo en muchas áreas cerebrales, incluida el área cingulada anterior, que se supone se activan al percibir dolor táctil. Algunas neuronas de esta área se activan también cuando el sujeto observa a otro siendo pinchado en la piel, lo que provoca una respuesta táctil esbozada en el propio sujeto cuando ve que otro es pinchado. Sentimos de modo esquemático en el mismo sitio de nuestra propia piel la sensación táctil del otro. Lo que parece evitar que la sensación sea tan intensa como la de algo real pinchándonos es la inhibición que producen nuestros nervios táctiles enviando señales hacia el córtex que son contradictorias con la que envían los sistemas visuales que ven el pinchazo. Sin embargo, Ramachandran (2011) informa de pacientes con síndrome de brazo fantasma, que sintieron que algo les pinchaba en su brazo inexistente, cuando vieron a otro siendo pinchado en su brazo. Aparentemente, la falta de señales procedentes del brazo fantasma, que pudieran contradecir a la percepción visual, permitió a las neuronas espejo del córtex sensorial del tacto generar una señal de contacto físico completo en la piel.
Según Ramachandran, en los cerebros humanos y sólo en estos, esta capacidad de las neuronas-espejo que ya existe en los monos se habría ampliado, de modo que cuando interpretamos intenciones complejas en las mentes de otros seres, habría neuronas espejo que se activarían en las áreas nuestras correspondientes a intenciones propias similares.
La anosognosia es un síndrome que tienen algunos pacientes que sufren de parálisis del lado izquierdo del cuerpo, por un trombo en el lado derecho, pero que niegan que sufran ninguna parálisis. Ramachandran comenta el sorprendente caso de algunos de estos pacientes que, cuando se les muestra otro paciente con parálisis en un lado del cuerpo, ellos niegan no sólo que ellos sufran parálisis, sino que el otro tenga tampoco parálisis alguna. Es como si el paciente, cada vez que trata de hacer un juicio sobre los movimientos de otra persona, tuviera que ejecutar “una simulación de realidad virtual de los movimientos correspondientes en su propio cerebro”. Pero como las neuronas espejo que simulan su propio movimiento del lado izquierdo están afectadas (de ahí que no perciba que su brazo no se mueve) no puede simular tampoco el movimiento o falta de movimiento del otro.
También sugiere que, en el aprendizaje, el núcleo dinámico en conjunción con el córtex prefrontal podría sistemáticamente inhibir todos los comportamientos motores imitativos que tienden a dispararse en la observación de la persona a imitar, y otros posibles que propondría el lóbulo parietal inferior, salvo el que sea más conveniente para alcanzar el objetivo asociado a la tarea.
Esta capacidad permitiría el imitar los movimientos de otro capaces de producir un artefacto o una nueva técnica, posibilitando así la herencia cultural. También permitiría imitar los movimientos de labios y lengua de otros, facilitando el aprendizaje del habla. De hecho, una de las regiones donde abundan las neuronas espejo según Rizzolatti (uno de sus descubridores) es en el área ventral premotora de los monos, una región que podría ser la precursora de nuestra área de Broca (el área del cerebro humano asociada con la producción del lenguaje sintáctico).
De hecho, “Cuando los circuitos inhibitorios frontales están dañados, como en el síndrome del lóbulo frontal, el paciente a veces imita gestos incontrolablemente, un síntoma llamado equopraxia.” Grados menores de pérdida de esta inhibición podrían explicar comportamientos cotidianos en adultos como los que apuntaba Charles Darwin: nos percibimos doblando inconscientemente la rodilla cuando vemos a un atleta a punto de lanzar la jabalina, y apretamos y aflojamos las mandíbulas cuando vemos a alguien usando un par de tijeras.

Orangután de Borneo usando una lanza para pescar
Los monos son capaces de imitar a otro mono en el uso de una piedra o un palo, pero sólo en el contexto de hacerse con comida. Un bebé, sin embargo, parece encontrar gran satisfacción en el mero hecho de imitar o ser imitado (Meltzoff y Marshall, 2018) y esto parece haber sido clave para el desarrollo de las culturas.
Referencias
Catmur, C., Gillmeister, H., Bird, G., Liepelt, R., Brass, M., & Heyes, C. (2008). Through the looking glass: counter-mirror activation following incompatible sensorimotor learning. European Journal of Neuroscience, 28(6), 1208–1215. doi: 10.1111/j.1460-9568.2008.06419.x
Dennett, Daniel C . (2017). From bacteria to Bach and back: the evolution of minds. W. W. Norton & Company.
Lakoff, G., Johnson M. (1999). Philosophy in the flesh. The embodied mind and its challenge to western thought. Basic Books, New York.
Leslie, A. (1997). The origins of Theory of Mid. Psychological Review. 94, 84-106.
Marr, D. (1985). La Visión, Alianza Psicología, Madrid.
Meltzoff, Andrew N. and Moore, M. Keith (1997). Explaining Facial Imitation: A Theoretical Model. Early Development and Parenting, Vol. 6, 179-192.
Meltzoff, A. N. and Marshall, P. J. (2018). Human infant imitation as a social survival circuit. Current Opinion in Behavioral Sciences, 24:130–136.
Ramachandran, V. S. (2011). The Tell-Tale Brain. W. W. Norton & Company.

Deja un comentario